Evento de extinción del Capitaniense
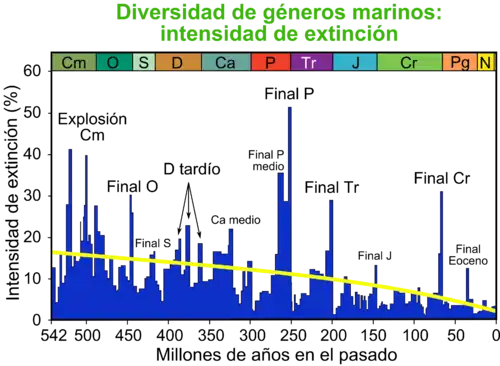
El evento de extinción del Capitaniense, también llamado extinción del final del Guadalupiense[1] o extinción masiva del Guadalupiense–Lopingiense,[2] tuvo lugar hace aproximadamente 262–259 Ma al final de la edad Capitaniense del Guadalupiense. Se considera la tercera gran crisis biótica del Fanerozoico, tras las extinciones ordovícica tardía y del Pérmico–Triásico.[3] Durante este intervalo hubo dos pulsos principales separados por casi 3 Ma; el primero en torno a 262 Ma y el segundo cerca de 259 Ma, ambos vinculados al vulcanismo de los traps de Emeishan, cambios climáticos extremos, anoxia oceánica y acidificación masiva.[4] Se estima que desapareció entre el 62 % y el 87 % de los géneros marinos, incluidos fusulínidos como Yabeina y Pseudoschwagerina, braquiópodos de los órdenes Orthida y Productida, corales tabulados como Halysites y algas calcáreas de los Dasycladales.[5] Registros en China, Sudáfrica, Spitsbergen, Canadá y la Cordillera Ibérica confirman que afectó a todo el planeta.[6]
Identificación
El límite Capitaniense–Wuchiapingiense se define por la primera aparición del conodonto Clarkina postbitteri postbitteri en el perfil de Penglaitan en Guangxi, China, datado en 259,51 ± 0,21 Ma mediante U–Pb en circones.[7] Lutitas negras de la Formación Maokou en Sichuan y de la Formación Madoupo en Guizhou muestran picos de mercurio de hasta 50 ng/g y anomalías de δ¹³C de −4 ‰, indicadores de los pulsos volcánicos y de perturbaciones del ciclo del carbono.[8] Biomarcadores como allerano y gammacerano y capas ricas en materia orgánica documentan dos episodios de anoxia oceánica (OAE-C1 y OAE-C2) en depósitos de Tetis occidental y Yangtsé.[9] Fósiles muestran extinciones masivas de fusulínidos (Schwagerina), corales rugosos (Lithostrotion) y braquiópodos (Spiriferida) en China y Tetis occidental; en tierra, desaparecieron gran parte de los Dinocephalia (Diictodon) y Anomodontia.[10]
Causas
El principal motor fue el gran magmatismo de los traps de Emeishan, que liberó más de 106 km³ de basaltos y entre 130 y 188 teratoneladas de CO₂ y SO₂. Esto elevó la temperatura media global hasta 5 °C y provocó lluvias ácidas que disolvieron carbonatos someros.[11] La estratificación de la columna de agua, impulsada por metales pesados y nutrientes volcánicos, generó euxinia y anoxia persistente en lutitas negras.[12] Fluctuaciones eustáticas con caídas del nivel del mar de decenas de metros colapsaron plataformas carbonatadas y arrecifes de Halysites y Waagenophyllidae.[13] Un impacto meteórico se descarta por la falta de cráteres sincrónicos o esférulas de impacto.[14] La configuración de Pangea restringió la ventilación oceánica y amplió zonas hipóxicas en latitudes tropicales, agravando la mortalidad selectiva.[12]
Consecuencias
El primer pulso, cerca de 262 Ma, eliminó hasta el 70 % de los fusulínidos como Yabeina y Polydiexodina y de braquiópodos como Spiriferida en Tetis occidental.[5] El pulso principal, alrededor de 259 Ma y coincidiendo con el pico de mercurio y la máxima erupción de los traps de Emeishan, provocó la desaparición del 87 % de braquiópodos en Spitsbergen y del 82 % de corales tabulados en Hungría.[12] La extinción de amonites como Timorites y Strigogoniatites alcanzó el 65 % de los géneros europeos y boreales.[12] Globalmente, la pérdida de biodiversidad marina osciló entre el 62 % y el 65 % de los géneros.[3] En tierra, los Dinocephalia (Diictodon feliceps) quedaron reducidos a un solo género y los Anomodontia a la mitad de su diversidad previa.[9] Los ambientes someros sufrieron pérdidas de hasta el 87 %, frente al 50 % en depósitos profundos (Borup Fiord).[12] Las latitudes bajas mostraron mortalidades superiores al 80 %, mientras que en latitudes altas existieron refugios parciales con menos del 50 % de pérdida.[3] Taxones calcificantes como fusulínidos, Rugosa y Dasycladales presentaron mayor selectividad que bivalvos, pteriomorfos y amonites oportunistas.[5] El colapso sincrónico en Tetis, Panthalassa y la Pangea Ártica reafirma su alcance global.[4] La diversidad de Orthida y fusulínidos de la familia Verbeekinidae cayó un 82 %, mientras que Timorites sobrevivió mejor con un 35 % de géneros remanentes.[5]
Recuperación
Menos de 1 Ma después, fosiles de fusulínidos oportunistas como Afghanella y bivalvos reaparecen en sedimentos de Madoupo.[12] Entre 2 y 5 Ma tras el evento, los arrecifes coralinos se reconstituyen, dominados por corales tabulados como Phaceloidea y briozoos de la familia Fenestellidae.[12] En tierra, dicinodontos como Diictodon y rizodontos ocuparon nichos vacantes, mientras que los terápsidos más especializados desaparecieron definitivamente.[9] Durante el Lopingiense y el Carniense se registra radiación de ammonoideos como Dzhulfites y crinoideos, señal de recuperación de ecosistemas pelágicos complejos.[7] La estabilización de ciclos biogeoquímicos y la diversificación post-crisis sentaron las bases para la biodiversidad triásica temprana.[12]
Véase también
- Extinción masiva del Pérmico-Triásico
- Pérmico
- Capitaniense
- Provincia ígnea de Emeishan
Referencias
- ↑ De la Horra, R.; Galán-Abellán, A. B.; López-Gómez, José; Sheldon, Nathan D.; Barrenechea, J. F.; Luque, F. J.; Arche, A.; Benito, M. I. (August–September 2012). «Paleoecological and paleoenvironmental changes during the continental Middle–Late Permian transition at the SE Iberian Ranges, Spain». Global and Planetary Change. 94-95: 46-61. Bibcode:2012GPC....94...46D. doi:10.1016/j.gloplacha.2012.06.008. Consultado el 15 de diciembre de 2022.
- ↑ Huang, Yuangeng; Chen, Zhong-Qiang; Zhao, Laishi; Stanley Jr., George D.; Yan, Jiaxin; Pei, Yu; Yang, Wanrong; Huang, Junhua (1 de abril de 2019). «Restoration of reef ecosystems following the Guadalupian–Lopingian boundary mass extinction: Evidence from the Laibin area, South China». Palaeogeography, Palaeoclimatology, Palaeoecology 519: 8-22. Bibcode:2019PPP...519....8H. S2CID 134096639. doi:10.1016/j.palaeo.2017.08.027. Consultado el 8 de enero de 2023.
- ↑ a b c «Mass extinction event 260 million years ago resulted from climate change, studies say». Ars Technica. 25 de julio de 2023. Consultado el 13 de mayo de 2025.
- ↑ a b Yang et al. (2024). «Mercury evidence of Emeishan volcanism driving the mid-Capitanian extinction». GSA Bulletin 136. doi:10.1130/B37796.1. Consultado el 13 de mayo de 2025.
- ↑ a b c d Clapham, Shen & Bottjer (2009). «Reassessing the severity and causes of the end-Guadalupian biotic crisis». Paleobiology 35: 32-50. Consultado el 13 de mayo de 2025.
- ↑ Song et al. (2023). «Global oceanic anoxia linked with the Capitanian marine mass extinction». Earth and Planetary Science Letters 610. doi:10.1016/j.epsl.2023.118128. Consultado el 13 de mayo de 2025.
- ↑ a b Jin et al. (2006). «Global Stratotype Section and Point for the boundary between the Capitanian and Wuchiapingian Stage (Permian)». Episodes 29 (4): 255-264. Consultado el 13 de mayo de 2025.
- ↑ Sun et al. (2017). «Mercury evidence from the Sino-Korean block for Emeishan volcanism during the Capitanian mass extinction». Geological Magazine 156: 931-944. Consultado el 13 de mayo de 2025.
- ↑ a b c Day & Rubidge (2021). «The Late Capitanian mass extinction of terrestrial vertebrates in the Karoo Basin of South Africa». Frontiers in Earth Science 9. Consultado el 13 de mayo de 2025.
- ↑ Tong et al. (2019). e0213456. «Capitanian mass extinction anoxia biomarkers». PLOS ONE 14. Consultado el 13 de mayo de 2025.
- ↑ Wignall et al. (2009). «Coupled volcanic activity and marine anoxia in the Upper Yangtze». Chemical Geology 263: 247-258. Consultado el 13 de mayo de 2025.
- ↑ a b c d e f g h Song et al. (2023). «Global oceanic anoxia linked with the Capitanian marine mass extinction». Earth and Planetary Science Letters 610. doi:10.1016/j.epsl.2023.118128. Consultado el 13 de mayo de 2025.
- ↑ Wignall & Twitchett (2002). «Mass extinctions and sea-level changes». Journal of the Geological Society 159 (4): 667-676. Consultado el 13 de mayo de 2025.
- ↑ «Capitanian Stage». Encyclopedia Britannica. 2025. Consultado el 13 de mayo de 2025.
Bibliografía
- Bond, D.P.G.; Wignall, P.B. (2009). The Capitanian (Middle Permian) mass extinction in China and its relationship to volcanic eruptions and climate change. Geological Journal.
- Shen, S.-Z. et al. (2019). A sudden end-Permian mass extinction in South China. Science Advances.
- Yang, P. et al. (2024). Mercury evidence of Emeishan volcanism driving the mid-Capitanian extinction. GSA Bulletin.
- Tong, J. et al. (2019). Capitanian mass extinction anoxia biomarkers. PLOS ONE.
- Day, M.O.; Rubidge, B.S. (2021). The late Capitanian mass extinction of terrestrial vertebrates in the Karoo Basin of South Africa. Frontiers in Earth Science.
- Song, H.; Algeo, T.J. et al. (2023). Global oceanic anoxia linked with the Capitanian marine mass extinction. Earth and Planetary Science Letters.
- Sun, Y. et al. (2017). Mercury evidence from the Sino-Korean block for Emeishan volcanism during the Capitanian mass extinction. Geological Magazine.
- Jin, Y.-G.; Shi, G.R. (1996). Permian extinction of marine invertebrates in Spitsbergen. Polar Research.
- Hall, R. et al. (2023). Astronomical forces as a potential cause for the end-Guadalupian transition. Earth-Science Reviews.
- Wignall, P.B.; Twitchett, R.J. (2002). The mid-Permian (Capitanian) mass extinction. Journal of the Geological Society.
Enlaces externos
- Esta obra contiene una traducción derivada de «Capitanian mass extinction event» de Wikipedia en inglés, concretamente de esta versión, publicada por sus editores bajo la Licencia de documentación libre de GNU y la Licencia Creative Commons Atribución-CompartirIgual 4.0 Internacional.
| Control de autoridades |
|
|---|
 Datos: Q21014121
Datos: Q21014121